

We are accustomed to thinking of inheritance as a clean, predictable distribution of structural assets — the crisp geometry of a Mendelian matrix. We inherit the bridge of a grandfather's nose, the deep melanin concentration of a maternal aunt's irises, or a clear genetic vulnerability to early-onset physiological conditions. We are taught by classical biology that our blueprint is written in an indelible, structural script — the double helix of deoxyribonucleic acid (DNA) — bequeathed to us at the absolute moment of conception, sterile and completely indifferent to the historical trials, geopolitical horrors, or physiological agonies of those who bore it before us. But if you look closely at the modern metabolic landscape of the Indian subcontinent, a profound and deeply unsettling medical anomaly emerges: a sweeping, unprecedented epidemic of type 2 diabetes, central adiposity, and premature ischemic heart disease that aggressively strikes individuals who have never known a single day of caloric starvation in their lives.
This distinct vulnerability cannot be explained by standard genetic variations alone; it defies classical evolutionary mutation rates, which operate across millennia rather than mere decades. To understand it, we are forced to look back past our parents, deeper into the dark, historical corridors of systemic trauma — specifically, to the bellies of our grandmothers (our Nanis and Dadis) who survived the cataclysmic, man-made famines of the British Raj. The core thesis of modern environmental epigenetics is both scientifically revolutionary and emotionally devastating: the human body remembers what history tries to clean from the ledger. When your maternal grandmother stood in an endless, agonizing ration queue in war-torn Bengal in 1943, or endured the silent, unyielding deprivation of late-nineteenth-century colonial resource drainage, her body was not merely suffering; it was actively computing. Faced with sudden, catastrophic macro-nutritional starvation, her physiology made a series of high-stakes metabolic calculations. It assumed, with absolute evolutionary logic, that the world she inhabited was one of permanent, unyielding, and brutal scarcity. To preserve the immediate survival of the fetus she might carry, and the nascent germlines already nested within that fetus, her body systematically altered the regulatory software governing her genome. It placed microscopic molecular "tags" — specifically methyl groups — onto strategic DNA sequences, winding specific genetic clusters tightly around structural proteins called histones to permanently silence them, while leaving others completely exposed. This was the rapid activation of the "thrifty phenotype," a sophisticated biological strategy designed to hoard lipids, restrict insulin sensitivity, and minimize caloric expenditure at all costs. Crucially, this software update did not dissipate when the famine broke. It was not wiped clean by the arrival of post-colonial development, national independence, or the relative abundance of modern supermarkets. Instead, through a complex process known as transgenerational epigenetic inheritance (TEI), these molecular scars were passed down the lineage, bypassing the standard cellular reprogramming that occurs during early embryonic development. Your body behaves as though it is still expecting the British grain ships to vanish from the horizon. When this historical software — meticulously optimized for extreme starvation — is dropped into a modern environment of caloric abundance and processed carbohydrates, the result is a catastrophic metabolic mismatch. Your Nani's famine is not a distant historical footnote; it is an active, biochemical reality operating inside your cells right now.
Section I: The Historical Crucible — Colonial Architecture of Starvation
To fully comprehend the biological mechanisms of inherited hunger, we must first fearlessly dismantle the historical forces that engineered it. The catastrophic famines that plagued the Indian subcontinent under British colonial rule were not simple, unavoidable acts of nature, nor were they the straightforward consequence of monsoon failures. Rather, they were highly structured, administrative triumphs of absolute laissez-faire free-market capitalism, predatory taxation, and imperial resource prioritization. From the Great Bengal Famine of 1770, which decimated a full third of the region's population under the rapacious revenue collection of the East India Company, to the devastating, late-Victorian famines documented by early economic thinkers like Dadabhai Naoroji, starvation was systematically normalized as an economic variable.
The absolute zenith of this administrative violence occurred during the Great Bengal Famine of 1943, where between 2.1 and 3 million human lives were utterly extinguished. Under the wartime administration of Prime Minister Winston Churchill and the implementation of the ruthless "denial policy," vast quantities of grain were systematically exported out of food-deficit Bengal to stock domestic European reserves and feed military personnel in the Mediterranean theatre. Simultaneously, local transport vessels, riverboats, and grain stocks were destroyed or confiscated to prevent potential Japanese advance, effectively paralyzing the internal distribution network of the delta. Rice prices skyrocketed exponentially, far out of reach for the rural proletariat — landless laborers, artisanal fishermen, and peasant farmers. This was what the Nobel laureate Amartya Sen famously categorized as an "entitlement failure." The food physically existed within the province, but the structural, violent mechanics of the colonial market rendered it biologically inaccessible to the populace.
"The starvation of millions was not an act of God, but an act of policy. The grain warehouses of Calcutta were full, yet the countryside was transformed into an open-air morgue. The physiological stress borne by the women of that era was an absolute, systemic compression of human biology."
For a pregnant woman surviving in this apocalyptic landscape, or a young prepubertal girl whose ovaries were already developing the oocytes that would one day become her children, the internal cellular environment was one of absolute, unmitigated crisis. Total daily caloric intake plummeted from a baseline of subsistence to less than 700 calories, composed primarily of nutrient-deficient wild roots, water, and boiled husks. This catastrophic macroeconomic deprivation was accompanied by a total collapse of essential micronutrients — folate, cobalamin (B12), zinc, and vital amino acids. It is precisely this lethal combination of severe caloric starvation and acute micronutrient depletion that serves as the perfect chemical catalyst for permanent epigenetic restructuring.
Section II: The Mechanics of Epigenetics — Beyond the Static Code
For nearly a century, the central dogma of molecular biology asserted that the flow of genetic information was completely unidirectional and entirely impervious to environmental intervention: DNA is transcribed into RNA, which is subsequently translated into functional proteins. Jean-Baptiste Lamarck's historical assertion that an organism could pass on characteristics acquired during its direct lifetime was dismissed as unscientific heresy. However, the emergence of contemporary molecular epigenetics has revealed a secondary, immensely complex layer of regulation that sits "above" the structural DNA sequence.
The DNA inside a single human cell is roughly two meters long, packed into a microscopic nucleus by wrapping around octameric protein complexes called histones. This architectural configuration is highly dynamic. If the DNA is wound too tightly around these histone cores (heterochromatin), the transcriptional machinery of the cell — such as RNA polymerase II and its associated transcription factors — cannot physically access the promoter regions of specific genes. The gene is effectively silenced. Conversely, if the chromatin architecture is relaxed (euchromatin), the gene can be actively transcribed. The molecular levers that control this architectural shifting are primarily three-fold.
1. DNA Methylation
This is the most stable and thoroughly studied epigenetic mechanism. It involves the covalent attachment of a methyl group to the 5-carbon position of a cytosine ring, typically within cytosine-guanine dinucleotide (CpG) islands located in or near gene promoters. This enzymatic reaction is catalyzed by DNA methyltransferases (DNMT1, DNMT3a, and DNMT3b). When a promoter region undergoes hypermethylation, it physically blocks transcription factor binding and recruits methyl-CpG-binding domain proteins (MBDs), which further compact the chromatin, turning the gene completely "off." This precise biochemical pathway is profoundly dependent on immediate nutrition. The universal methyl donor for this reaction is S-adenosylmethionine (SAM), which is synthesized via the one-carbon metabolism pathway. This pathway relies entirely on dietary intakes of folate, vitamin B12, choline, and methionine. When a pregnant woman faces severe famine, the total shortage of these micronutrients severely disrupts her one-carbon metabolism, leading to widespread hypomethylation or inappropriate hypermethylation across her fetus's developing genome.
2. Histone Modifications
The amino-terminal tails of histone proteins extend outward from the core nucleosome and are subject to an array of post-translational modifications, including acetylation, methylation, phosphorylation, and ubiquitination. For instance, histone acetylation — catalyzed by histone acetyl transferases (HATs) — neutralizes the positive charge of lysine residues, weakening the electrostatic bond between the histone and the negatively charged DNA backbone. This opens the chromatin structure, permitting active transcription. Histone deacetylases (HDACs), conversely, remove these acetyl groups, sealing the DNA away from transcription. The activity of these enzymes is directly regulated by cellular metabolic intermediates such as acetyl-CoA and NAD+, linking maternal nutritional status directly to chromatin structure.
3. Non-Coding RNAs (ncRNAs)
Micro RNAs (miRNAs) and long non-coding RNAs (lncRNAs) do not code for proteins themselves, but act as post-transcriptional regulators. They bind to complementary sequences on messenger RNA (mRNA) transcripts, causing them to degrade or preventing them from being translated into proteins. Recent breakthroughs in reproductive biology demonstrate that these small non-coding RNAs are packaged into extracellular vesicles within the uterus and the male epididymis, serving as a rapid response system for transmitting environmental stress profiles across generations.
Section III: The Molecular Legacy — University and Clinical Evidence
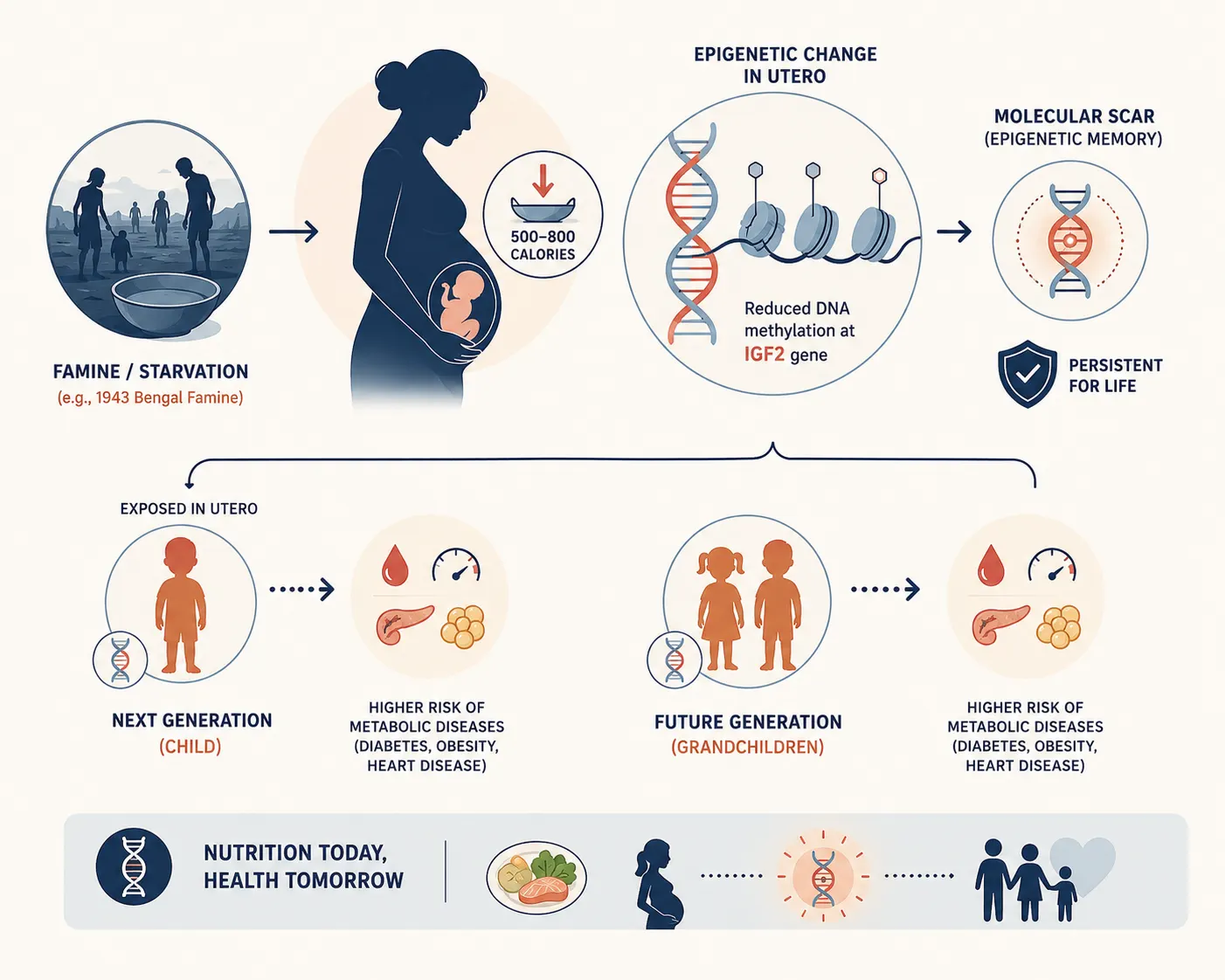
The foundational proof that maternal starvation leaves an indelible epigenetic signature on future generations comes from a series of extraordinary, long-term epidemiological and molecular studies conducted by research institutions worldwide. While direct tissue samples from the 1943 Bengal famine are tragically non-existent due to historical neglect, parallel cohorts — most notably the Dutch Hunger Winter and the Great Chinese Famine — have provided the precise empirical models that explain the South Asian metabolic crisis.
The Dutch Hunger Winter Cohort
Between November 1944 and April 1945, the Nazi occupation forces placed a total embargo on food transport in the western Netherlands. This created a highly circumscribed, sudden, and well-documented period of severe famine affecting an otherwise well-nourished population. Daily rations dropped to between 500 and 800 calories. Decades later, a team of researchers from Leiden University Medical Center and Columbia University, led by Dr. L.H. Lumey and Dr. Bas Heijmans, tracked down individuals who were conceived or carried in utero during this specific window. Their findings, published in landmark papers in The Lancet and the Proceedings of the National Academy of Sciences (PNAS), were revolutionary. Sixty years after the famine ended, individuals who were exposed to starvation in utero during early gestation displayed a distinct, persistent reduction in DNA methylation at the imprinted IGF2 (Insulin-like Growth Factor II) gene compared to their unexposed, same-sex siblings. The IGF2 gene is a master controller of fetal growth and human metabolic programming. This change in methylation was completely independent of the individuals' adult lifestyle or socioeconomic status; it was a permanent molecular scar left by their mother's winter of starvation.
Empirical Insight: The Leiden-Columbia Molecular Discovery
The 2008 study by Heijmans et al. demonstrated that early prenatal exposure to famine was associated with a significant decrease in the methylation of the IGF2 locus. This hypomethylation leads to the chronic, unregulated over-activation or under-activation of growth factors, laying the structural groundwork for metabolic syndrome, elevated LDL cholesterol, and an increased Body Mass Index (BMI) six decades later.
The Thrifty Phenotype Hypothesis and the Barker Legacy
To contextualize these findings, we must turn to the work of British epidemiologist Dr. David Barker. In the late 1980s, Barker formulated the "Fetal Origins of Adult Disease" hypothesis. Investigating geographical patterns of infant mortality and adult ischemic heart disease in the United Kingdom, Barker observed a profound paradox: areas with the highest rates of infant mortality in the early twentieth century also had the highest rates of adult coronary heart disease decades later. Barker argued that when a developing fetus is deprived of essential nutrients, it undergoes a permanent physiological reorganization. To maximize the chances of immediate survival, the fetus diverts its limited resources toward the development of its most vital organ — the brain — at the expense of peripheral organs like the pancreas, kidneys, and skeletal muscle. This results in a child born with a reduced number of insulin-producing pancreatic beta-cells, a lower nephron count in the kidneys, and an altered vascular architecture. If this child subsequently encounters an environment of nutritional security or surplus, their down-regulated metabolic machinery is completely overwhelmed. The "thrifty" body simply cannot handle the caloric load.
| Fetal Organ Affected | Structural / Physiological Modification | Adult Clinical Consequence |
|---|---|---|
| Endocrine Pancreas | Reduced volume and density of pancreatic beta-cells; impaired vascularization of islets. | Diminished insulin secretion capacity; early onset of Type 2 Diabetes Mellitus. |
| Skeletal Muscle | Decreased muscle fiber numbers; down-regulated GLUT4 glucose transporter expression. | Profoundly reduced peripheral glucose clearance; systemic insulin resistance. |
| Adipose Tissue | Shift toward visceral adipocyte distribution; altered adipokine secretion profile (Leptin/Adiponectin ratio). | Central/trunk obesity; chronic, low-grade systemic inflammation. |
| Kidneys / Renal System | Reduced total nephron count; compensatory hypertrophy of remaining glomeruli. | Systemic arterial hypertension; accelerated chronic kidney disease (CKD). |
| HPA Axis | Hyper-reactivity of the paraventricular nucleus; down-regulation of glucocorticoid receptors. | Chronic hypercortisolemia; impaired stress resilience; elevated visceral fat deposition. |
Section IV: The South Asian Paradox — A Twice-Traumatized Genome
The global medical community has long puzzled over the "South Asian Paradox." Individuals of Indian, Pakistani, and Bangladeshi descent exhibit a unique phenotypic profile characterized by a remarkably high percentage of body fat relative to body mass index (BMI), profound insulin resistance at low body weights, visceral fat deposition around the internal organs, and an extraordinarily high susceptibility to cardiovascular disease. A South Asian man with a BMI of 22 can display the exact same metabolic and diabetic risk profile as a Caucasian man with a BMI of 30.
For decades, public health campaigns lazily attributed this phenomenon to modern lifestyle changes, rising affluence, and a diet rich in refined carbohydrates and clarified butter (ghee). But this explanation ignores a critical epidemiological fact: the onset of metabolic disease in South Asians occurs a decade earlier than in Western populations, and it occurs across all socioeconomic strata, including impoverished rural communities. The answer does not lie merely in the modern kitchen; it lies in the historical timeline of the subcontinent's famines. Unlike the Dutch Hunger Winter, which was a single, isolated event lasting six months, the Indian subcontinent was subjected to an unprecedented series of massive famines over a span of two centuries under British rule. Between 1770 and 1947, India experienced over 30 major famines. This represents a sustained, multi-generational evolutionary pressure. In the context of evolutionary biology, a single generation of starvation triggers an intergenerational response (affecting the fetus directly inside the mother's womb). However, when a population is subjected to repeated, systemic nutritional deprivation over consecutive generations, the epigenetic adjustments consolidate into a true transgenerational program.
When your Nani was pregnant with your mother during a period of colonial deprivation, her body was not encountering a novel, temporary shock. Her own genome had already been pre-programmed by her mother's experiences during the late-Victorian holocausts. The South Asian genome has been compressed by history multiple times. It is a twice-traumatized biological archive. The epigenetic software has been locked into a permanent "crisis mode." When the post-independence era finally brought relative food security and the green revolution, it introduced an unprecedented influx of high-glycemic carbohydrates, refined seed oils, and a sedentary lifestyle. This created an extreme evolutionary mismatch. The "thrifty" phenotype, programmed to store every single calorie as visceral fat to survive the next inevitable colonial grain extraction, was suddenly submerged in an environment of constant caloric availability. The result is a metabolic explosion. The high prevalence of type 2 diabetes in South Asia today is the direct, biological bill of British imperial economic policies, coming due three generations later.
Section V: The HPA Axis and Chronic Cortisol Inundation
The inherited legacy of hunger is not restricted to how your body processes glucose or stores lipids; it fundamentally alters how your brain and endocrine systems perceive threat. At the absolute center of this physiological adaptation is the Hypothalamic-Pituitary-Adrenal (HPA) axis, the primary neuroendocrine system responsible for managing homeostatic stress responses.
Under normal physiological conditions, a perceived stressor triggers the paraventricular nucleus of the hypothalamus to secrete Corticotropin-Releasing Hormone (CRH). This hormone stimulates the anterior pituitary gland to release Adrenocorticotropic Hormone (ACTH), which travels through the bloodstream to the adrenal cortex, prompting the synthesis and systemic release of glucocorticoids, primarily cortisol. Cortisol prepares the organism for immediate survival: it elevates blood pressure, mobilizes glucose reserves via hepatic gluconeogenesis, and suppresses non-essential systems like digestion, reproduction, and immune function. Once the threat passes, cortisol binds to high-affinity Glucocorticoid Receptors (GR) in the hippocampus, activating a negative feedback loop that shuts down the entire cascade, returning the body to baseline.
However, when a pregnant mother experiences the chronic, existential trauma of a famine, her internal systemic cortisol levels remain permanently elevated. This flood of maternal cortisol overwhelms the placental enzyme 11-beta-hydroxysteroid dehydrogenase type 2 (11-beta-HSD2), which normally inactivates maternal cortisol into cortisone to protect the developing fetus. Consequently, the fetal brain is directly exposed to toxic levels of stress hormones during critical windows of neurodevelopment.
Research led by Dr. Rachel Yehuda at the Icahn School of Medicine at Mount Sinai, alongside collaborative global studies on historical trauma cohorts, shows that this in utero exposure leads to the permanent hypermethylation of the NR3C1 gene promoter in the fetal brain. The NR3C1 gene encodes the human glucocorticoid receptor. When this gene is hypermethylated and silenced, the child is born with an abnormally low density of glucocorticoid receptors in the hippocampus. The clinical consequence of this molecular alteration is profound. Because the negative feedback loop is broken, the individual's HPA axis is permanently hyper-reactive. They do not possess a normal, functioning psychological baseline. A minor everyday stressor — a financial worry, an interpersonal conflict, or workplace pressure — is interpreted by the neuroendocrine system as an imminent, life-threatening famine. The body responds by flooding the bloodstream with excess cortisol and adrenaline. This chronic hypercortisolemia directly drives visceral fat accumulation, elevates systemic arterial blood pressure, degrades insulin sensitivity in skeletal muscle, and maintains a state of low-grade vascular inflammation that accelerates coronary artery disease. Your Nani's terror of starvation has been translated into an ongoing, biochemical anxiety that shapes your modern psychological and physiological response to the world.
This inherited HPA hyper-reactivity is also a key driver behind why chronic occupational stress hits South Asian professionals harder at the metabolic level than population averages would predict — an intersection explored further in Why Peak Burnout Hits Indians at 25.
Section VI: Breaking the Cycle — Epigenetic Reversibility and Therapeutics
The realization that our bodies are living archives of historical trauma can feel like a form of biological fatalism. It is easy to interpret this data as a life sentence — an inescapable genetic destiny forged by dead administrators in colonial offices and borne helplessly by our ancestors. But this conclusion misses the most vital, redeeming feature of the epigenetic system: unlike the underlying DNA code, which is static, epigenetic modifications are inherently dynamic and potentially reversible.
The molecular tags placed upon our genes are meant to be responsive to the environment. Just as a period of profound trauma can write scars onto the chromatin architecture, targeted, conscious interventions can act as molecular erasers, rewriting the regulatory software of our cells. The emerging frontier of epigenetic therapeutics and lifestyle medicine offers a definitive path toward breaking this transgenerational cycle.
1. Nutritional Interventions and One-Carbon Support
Given that DNA methylation is directly dependent on the availability of methyl donors from our diet, targeted nutritional strategies can alter gene expression profiles. Diets rich in natural methyl donors and one-carbon metabolism cofactors — such as green leafy vegetables (folate), cruciferous vegetables (which contain sulforaphane, a potent HDAC inhibitor), and legumes — can assist in normalizing methylation levels at disrupted loci like IGF2. Furthermore, reducing the consumption of ultra-processed foods and high-glycemic index carbohydrates is absolutely critical for South Asians to prevent overwhelming a metabolic system programmed for thriftiness.
2. Physical Intervention and Chromatin Remodeling
Clinical studies conducted at institutions like Lund University have demonstrated that regular exercise induces rapid, widespread changes in DNA methylation patterns across human adipose and skeletal muscle tissue. Specifically, consistent resistance training and high-intensity interval training (HIIT) have been shown to decrease methylation at gene promoters responsible for glucose transport (GLUT4) and mitochondrial biogenesis (PGC-1α). This directly counteracts the inherited structural deficits in skeletal muscle capacity characteristic of the Barker phenotype, restoring peripheral insulin sensitivity.
3. Stress-Reduction and Somatic Reprogramming
Because the HPA axis in descendants of trauma is hyper-reactive due to the epigenetic silencing of the NR3C1 gene, interventions that actively down-regulate the sympathetic nervous system are powerful molecular interventions. Randomized controlled clinical trials have demonstrated that sustained mindfulness-based stress reduction (MBSR), deep pranayama breathing, and structured cognitive behavioral therapy can actually reverse the hypermethylation of the NR3C1 promoter. By consciously lowering systemic cortisol levels over time, we allow the hippocampal feedback loop to restore balance, reducing the neuroendocrine drive behind visceral fat accumulation and vascular inflammation.
Conclusion: The Living Archive
The science of transgenerational epigenetic inheritance forces us to completely re-evaluate the boundaries of the human individual. You are not a solitary biological actor operating in a historical vacuum. You are a continuation of a long, unbroken lineage of survival. The boundaries of your skin do not begin or end with your own birth certificate; they extend backward into the dust of colonial Bengal, through the dry, parched fields of the Deccan plateau, and into the crowded ration queues of a stolen empire. Your Nani's famine lives inside your body. It reveals itself in the way your cells process glucose, the way your liver synthesizes lipids, and the way your heart reacts to chronic stress. But this realization is not a sentence of inescapable disease; it is an invitation to profound somatic self-awareness. When you understand that your body's tendency to store fat or experience heightened stress is not a personal failure of willpower, but a sophisticated, inherited survival strategy engineered by your ancestors to keep you alive, your relationship with your health shifts from shame to deep reverence.
By making conscious choices — by eating with historical awareness, moving to remodel our muscle architecture, and practicing somatic healing to calm a hyper-reactive nervous system — we do more than just manage our personal health. We act as conscious molecular editors of our family's lineage. We honor the immense suffering of our grandmothers by ensuring that the survival programs they forged in the fires of historical tragedy are gracefully decommissioned, allowing future generations to inherit a biology defined not by the memory of hunger, but by the reality of freedom.
The same metabolic asymmetry discussed throughout this treatise — the body's deeply circadian relationship with food, stress, and hormonal timing — is explored through the lens of Ayurvedic science and modern chrononutrition in The Architecture of Time.
Read Further
-
Heijmans, B.T. et al. Persistent Epigenetic Differences Associated with Prenatal Exposure to Famine in Humans. Proceedings of the National Academy of Sciences (PNAS), 2008. — pubmed.ncbi.nlm.nih.gov/18955703
-
Barker, D.J. The Fetal and Infant Origins of Adult Disease. BMJ, 1990. — pmc.ncbi.nlm.nih.gov/articles/PMC1664286
-
Yehuda, R. et al. Holocaust Exposure Induced Intergenerational Effects on FKBP5 Methylation. Biological Psychiatry, 2015. — biologicalpsychiatryjournal.com
Disclaimer: The scientific mechanisms, epigenetic data, historical analyses, and clinical evidence presented in this article are synthesized from peer-reviewed research in molecular biology, epigenetics, endocrinology, and colonial economic history. This content is provided purely for educational and informational purposes and should not be construed as specific medical, dietary, or clinical advice. Consult a qualified physician before making any significant changes to your health regimen.